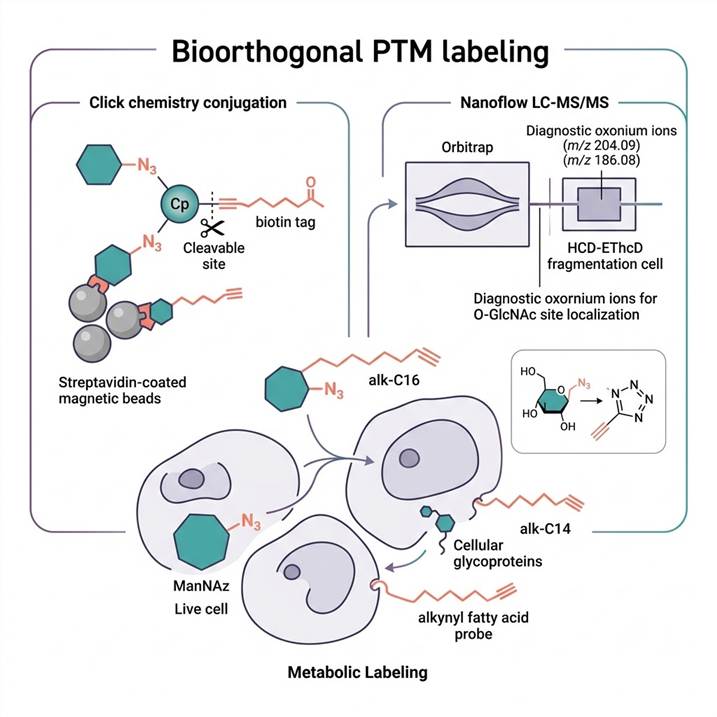
Chemical Selectivity for PTM Classes Beyond Antibody Reach
Post-translational modifications that involve substantial chemical remodeling of amino acid side chains — glycosylation (addition of complex glycans), lipidation (palmitoylation, myristoylation, prenylation), O-GlcNAcylation (single N-acetylglucosamine on Ser/Thr), and modifications lacking well-characterized antibodies — present a fundamental enrichment challenge for MS-based proteomics. Antibody-based immunoaffinity enrichment, the dominant strategy for phosphorylation, acetylation, and ubiquitination, is limited for these PTM classes by antibody availability, epitope masking by the modification itself, cross-reactivity with related glycan or lipid structures, and the inability to recognize the broad structural diversity of glycans and lipid modifications.
Bioorthogonal labeling bypasses these limitations entirely by introducing a chemically defined enrichment handle into the modified protein before capture. The strategy follows three universal steps: (1) metabolic or enzymatic incorporation of a bioorthogonal functional group (azide or alkyne) into target PTM substrates using modified sugar, lipid, or amino acid precursors; (2) chemoselective conjugation of the incorporated azide or alkyne with a complementary functional group on an affinity probe (typically biotin or FLAG tag) via click chemistry; (3) affinity capture and LC-MS identification of the labeled modification sites. Because the enrichment handle is chemically defined and orthogonal to native biological functional groups, this approach delivers specificity and reproducibility that is independent of antibody quality and can be applied to any PTM for which a bioorthogonal precursor can be metabolized by the cell or incorporated by an enzyme. For conventional enrichment approaches where antibodies are available and well-validated, our Modified Peptide Enrichment Services provide complementary immunoaffinity and metal-affinity options.
Find Your Solution: Research Goal → Bioorthogonal Labeling Strategy
| Your Research Goal |
Recommended Labeling Strategy |
Key Techniques |
| Profiling O-GlcNAc-modified proteins and site mapping in cell culture models |
Metabolic glycan labeling with GalNAz + CuAAC click chemistry |
GalNAz incorporation via salvage pathway, CuAAC with alkyne-biotin (cleavable linker), streptavidin enrichment, HCD-EThcD LC-MS/MS, HexNAc diagnostic neutral loss |
| Identifying S-palmitoylated proteins and site-specific mapping of S-acylation |
Metabolic labeling with alkynyl palmitate (alk-C16) + CuAAC click chemistry |
Alk-C16 incorporation, CuAAC biotin conjugation, streptavidin capture, hydroxylamine cleavage of thioester bond for mild elution, LC-MS/MS with site localization |
| Characterization of N-glycan and O-glycan occupancy and glycan heterogeneity |
Azido sugar labeling (ManNAz, GalNAz, GlcNAz) + click enrichment |
Metabolic incorporation of azido monosaccharides, click conjugation to cleavable biotin, glycopeptide enrichment, intact glycopeptide LC-MS/MS with HCD stepping fragmentation |
| Profiling N-myristoylated proteins in live cells or in vivo models |
Alkynyl myristate (alk-C14) metabolic labeling + CuAAC |
Alk-C14 metabolic incorporation by NMT, CuAAC biotinylation, streptavidin enrichment, tryptic digestion, LC-MS/MS with N-terminal myristoylation site confirmation |
| Tracking nascent (newly synthesized) protein modifications in pulse-chase experiments |
AHA (azidohomoalanine) or unnatural amino acid incorporation + click chemistry |
Methionine replacement with AHA, bioorthogonal tag conjugation, time-resolved enrichment of newly synthesized proteins, SILAC or TMT quantification for turnover dynamics |
| In vivo labeling of modified proteins in animal models or tissues |
SPAAC click chemistry with strained alkynes (DBCO-biotin) |
Metabolic probe administration (e.g., GalNAz in drinking water), SPAAC with DBCO-biotin for copper-free labeling, tissue homogenization, enrichment, LC-MS/MS |
Bioorthogonal Labeling Platforms for PTM Enrichment
Our service portfolio encompasses three complementary bioorthogonal labeling platforms, each optimized for specific PTM classes, sample types, and experimental contexts. The choice of labeling strategy determines the modification type that can be captured, the compatibility with live-cell versus lysate-based labeling, and the downstream LC-MS workflow required for site-specific identification.
Metabolic Glycan Labeling for O-GlcNAc and Glycoproteomics
Azido-modified monosaccharides — including N-azidoacetylgalactosamine (GalNAz), N-azidoacetylglucosamine (GlcNAz), and N-azidoacetylmannosamine (ManNAz, metabolic precursor to sialic acid) — are incorporated into cellular glycoproteins and O-GlcNAc-modified proteins through endogenous metabolic pathways. GalNAz is converted to UDP-GalNAz and incorporated by O-GlcNAc transferase (OGT) directly into O-GlcNAc-modified Ser/Thr residues, while also entering the sialic acid biosynthetic pathway for glycan labeling. After metabolic incorporation (typically 24–72 hours in cell culture), cell lysates are subjected to CuAAC click chemistry with an alkyne-functionalized cleavable biotin tag. Biotinylated proteins are enriched on streptavidin beads, and labeled peptides are released by cleavage of the acid-labile or photocleavable linker for LC-MS/MS analysis. HCD product-dependent EThcD (HCD-pd-EThcD) fragmentation is used for unambiguous localization of O-GlcNAc sites. For comprehensive glycoproteomics analysis beyond bioorthogonal labeling, our Glycoproteomics Analysis Services provide complementary lectin-based and HILIC-based enrichment platforms.
Alkynyl Fatty Acid Labeling for Protein Lipidation (Palmitoylation, Myristoylation)
ω-Alkynyl fatty acid analogs — including 16-alkynyl hexadecanoic acid (alk-C16 for palmitoylation), 14-alkynyl tetradecanoic acid (alk-C14 for myristoylation), and 18-alkynyl octadecanoic acid (alk-C18 for stearoylation) — are metabolically incorporated into cellular proteins by endogenous acyltransferases. After incorporation (typically 4–16 hours), labeled proteins are conjugated via CuAAC to azido-biotin probes with a cleavable linker (hydroxylamine-cleavable for S-acylation thioester bonds, or acid-cleavable for amide-linked myristoylation). Enriched proteins are identified by LC-MS/MS, with site localization achieved through the detection of modified cysteine (S-palmitoylation, loss of thioester bond upon hydroxylamine treatment) or N-terminal glycine (N-myristoylation). The hydroxylamine cleavage step itself provides orthogonal validation, as only thioester-linked S-acyl modifications are released, while amide-linked myristoylation remains bead-bound. For dedicated lipidation-focused investigations, our Protein Lipidation Analysis and Palmitoylation Analysis services provide specialized MS-based characterization of lipid-modified proteins.
Unnatural Amino Acid Labeling for Nascent Modifications and Pulse-Chase Proteomics
Bioorthogonal labeling of newly synthesized proteins is achieved through metabolic replacement of methionine with azidohomoalanine (AHA) in methionine-free culture media. Following a defined pulse period (15–120 minutes), AHA-incorporated nascent proteins are conjugated to alkyne-biotin via CuAAC and enriched for LC-MS/MS identification. When combined with SILAC or TMT quantification in a pulsed-SILAC or BONCAT (bioorthogonal non-canonical amino acid tagging) experimental design, this approach enables time-resolved measurement of protein and modification turnover dynamics. For studies of nascent PTMs specifically — such as monitoring the appearance of glycosylation, phosphorylation, or acetylation on newly synthesized proteins — the AHA labeling pulse can be coupled with parallel enrichment of the modification of interest. This dual-enrichment strategy is particularly powerful for dissecting the temporal sequence of co-translational and early post-translational modification events.
Why Choose Our Bioorthogonal PTM Labeling Services
Chemistry-Grade Specificity Independent of Antibody Availability
Unlike immunoaffinity methods that depend on antibody quality, lot consistency, and epitope accessibility, bioorthogonal labeling achieves enrichment through chemoselective click chemistry between an incorporated azide/alkyne and its complementary reaction partner. The specificity is defined by the metabolic or enzymatic incorporation step, not by antibody binding — enabling enrichment of PTM classes, glycan structures, and lipid modifications for which no suitable antibodies exist.
Multi-Probe Platform Under One Roof
Our platform spans azido sugars (GalNAz, GlcNAz, ManNAz), alkynyl fatty acids (alk-C14, alk-C16, alk-C18), and unnatural amino acids (AHA), covering O-GlcNAc, N-glycan, O-glycan, sialic acid, palmitoylation, myristoylation, stearoylation, and nascent protein labeling. This breadth enables integrated multi-PTM projects — for example, comparing O-GlcNAc and S-palmitoylation dynamics in the same signaling pathway — within a single service relationship.
Cleavable Linker Technology for Clean Release and Maximum Identification
All click chemistry enrichment steps use cleavable biotin linkers (acid-labile, photocleavable, or hydroxylamine-cleavable) rather than on-bead digestion. This approach eliminates streptavidin peptide contamination in MS samples, enables efficient recovery of labeled peptides, and allows the released peptides to be analyzed by standard LC-MS workflows without the interference of large amounts of bead-derived material in the digest.
Site-Specific Modification Localization with Advanced Fragmentation
For labile modifications such as O-GlcNAc and S-palmitoylation, standard HCD fragmentation often causes modification loss before peptide backbone cleavage, precluding site localization. We employ HCD product-dependent EThcD (HCD-pd-EThcD) and stepped HCD fragmentation strategies that preserve modification information while providing comprehensive sequence coverage for unambiguous site assignment — delivering site-level modification identification, not just protein-level labeling confirmation.
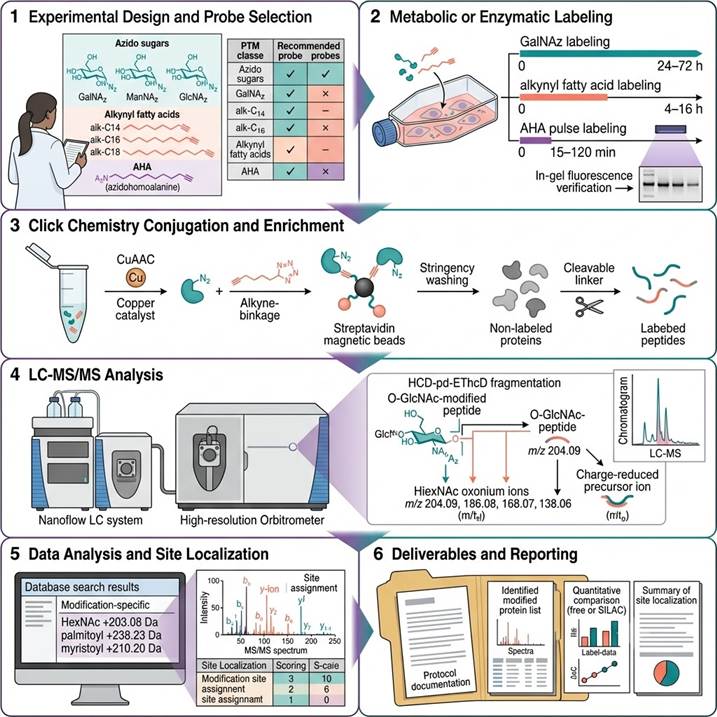
Workflow: From Probe Incorporation to Site-Level Modification Identification
Step 1: Experimental Design and Probe Selection
We consult with you to select the optimal bioorthogonal labeling strategy — azido sugar, alkynyl fatty acid, or AHA — based on the target PTM class, cell type or model system, labeling duration, and compatibility with downstream lysis and enrichment protocols. Metabolic labeling conditions (probe concentration, incubation time, serum concentration) are optimized for your specific cell or tissue system.
Step 2: Metabolic or Enzymatic Labeling
Bioorthogonal probes are introduced into the experimental system: GalNAz/GlcNAz/ManNAz for glycan and O-GlcNAc labeling (24–72 h, 50–200 µM), alkynyl fatty acids for lipidation labeling (4–16 h, 10–100 µM), or AHA for nascent protein labeling (15–120 min pulse, 1–4 mM in Met-free media). Labeling efficiency is verified by in-gel fluorescence (azide-alkyne ligation with fluorescent alkyne/azide) before proceeding to enrichment.
Step 3: Click Chemistry Conjugation and Enrichment
Cell lysates or tissue homogenates are subjected to CuAAC or SPAAC click chemistry with the complementary affinity probe (alkyne-biotin or azido-biotin with cleavable linker). Biotinylated proteins are captured on streptavidin magnetic beads, stringently washed, and specifically released by cleavable linker cleavage — avoiding on-bead digestion and minimizing MS interference.
Step 4: LC-MS/MS Analysis
Released peptides are analyzed by nanoflow LC-MS/MS on high-resolution Orbitrap platforms. For O-GlcNAc-modified peptides, HCD-pd-EThcD fragmentation is employed for unambiguous site localization. For palmitoylation, hydroxylamine cleavage specificity provides orthogonal validation. Intact glycopeptide analysis uses stepped HCD fragmentation for glycan and peptide sequence information acquisition.
Step 5: Data Analysis and Site Localization
LC-MS/MS data are searched against protein sequence databases with modification-specific parameters: HexNAc (+203.08 Da) on Ser/Thr for O-GlcNAc, S-palmitoylation (+238.23 Da) on Cys, N-myristoylation (+210.20 Da) on N-terminal Gly, and variable glycan compositions for glycoproteomics. Site localization confidence is assessed by modification-specific fragment ions and diagnostic neutral loss patterns.
Step 6: Deliverables and Reporting
Comprehensive report including: labeling and enrichment protocol documentation, LC-MS/MS acquisition parameters and data quality metrics, identified modified protein lists with site localization scores and spectra, quantitative comparison across experimental groups (label-free or SILAC), and annotated MS/MS spectra for each identified modification site.
Applications in Bioorthogonal PTM Labeling and Enrichment Proteomics
Bioorthogonal labeling is the method of choice for PTM classes and experimental contexts where antibody-based enrichment is impractical, unavailable, or insufficiently specific. The following application areas represent the most common and impactful uses of this technology.
O-GlcNAc Proteome Mapping in Disease Models
O-GlcNAcylation is a dynamic, nutrient-responsive modification that regulates transcription, metabolism, and signaling. Unlike phosphorylation, O-GlcNAc lacks a universal enrichment antibody with sufficient specificity and coverage for deep proteomic profiling. GalNAz metabolic labeling followed by click chemistry enrichment has become the gold-standard approach for O-GlcNAc proteomics, routinely identifying 500–2,000 O-GlcNAc sites per experiment. Our service applies this strategy to disease-relevant cell models, including cancer cell lines, stem cells, and primary neurons, with SILAC-based quantification for comparative O-GlcNAc profiling across drug treatment, metabolic perturbation, or disease-state conditions. For established O-GlcNAc targets requiring targeted verification, our Ultra-Sensitive Modified Protein Detection service provides complementary MRM/PRM-based validation.
Protein S-Palmitoylation and S-Acylation Profiling
Protein S-palmitoylation — the reversible addition of C16:0 palmitate to cysteine residues via a thioester bond — regulates protein membrane tethering, trafficking, and signaling complex assembly. Alkynyl palmitate (alk-C16) metabolic labeling coupled with CuAAC click enrichment is the most widely adopted approach for palmitoyl-proteomics, enabling identification of hundreds to thousands of palmitoylated proteins per experiment. The hydroxylamine-labile thioester bond provides an additional layer of specificity: only S-acylated proteins are released from streptavidin beads upon hydroxylamine treatment, while N-myristoylated and other non-thioester lipidated proteins remain bead-bound. This differential elution strategy can be exploited for parallel profiling of both S-acylated and N-myristoylated proteomes from a single alk-C14/alk-C16 labeling experiment.
N-Glycan and O-Glycan Occupancy and Heterogeneity Analysis
Glycosylation is the most structurally diverse PTM, presenting a formidable enrichment challenge for MS-based glycoproteomics. Azido sugar metabolic labeling (ManNAz for sialylated glycans, GalNAz for O-glycans, GlcNAz for N-glycans) enables selective enrichment of glycoproteins through a chemically defined handle that is independent of glycan structure, branching complexity, or sialylation status. After click conjugation to cleavable biotin and streptavidin enrichment, intact glycopeptides are analyzed by stepped HCD fragmentation that acquires both glycan oxonium ions (low m/z) and peptide backbone fragments (high m/z) in a single analysis. This approach provides simultaneous identification of peptide sequence, glycosylation site, and glycan composition for each detected glycopeptide. For dedicated glycan structural characterization, our Glycoproteomics Analysis Services provide complementary enzymatic and chemical deglycosylation workflows.
N-Myristoylation Profiling in Live Cells
N-myristoylation — the irreversible attachment of C14:0 myristate to N-terminal glycine residues by N-myristoyltransferase (NMT) — is a critical co-translational modification that regulates protein membrane targeting and function. Alkynyl myristate (alk-C14) metabolic labeling, combined with CuAAC click enrichment and LC-MS/MS, enables proteome-wide identification of N-myristoylated proteins with site-specific confirmation through detection of N-terminal myristoylated glycine peptides. This approach is particularly valuable for identifying NMT substrates in pathogens (including malaria, HIV, and trypanosomatid parasites) where NMT is being pursued as a therapeutic target. For broader lipidation analysis across different lipid classes, our Protein Lipidation Analysis service provides comprehensive MS-based lipidated protein characterization.
Nascent Proteome and Modification Turnover Dynamics
Bioorthogonal non-canonical amino acid tagging (BONCAT) using AHA (azidohomoalanine) enables time-resolved enrichment and identification of newly synthesized proteins. When combined with PTM-specific enrichment — for example, AHA labeling followed by parallel phosphopeptide enrichment or O-GlcNAc enrichment — this approach reveals the temporal dynamics of modification acquisition on nascent proteins. Pulse-chase AHA labeling with SILAC or TMT quantification provides turnover rate information for individual modification sites, distinguishing stable modifications from rapidly cycling ones. This capability is essential for understanding modification dynamics in signaling pathways, circadian regulation, and drug-response mechanisms where modification timing is as informative as modification identity.
Case Study: Metabolic Glycan Labeling for O-GlcNAc Proteome Profiling in Rice — Identification of 1,591 O-GlcNAcylation Sites Across the Plant Proteome Using GalNAz Labeling and Click Chemistry Enrichment
In a 2023 study published in Plant Biotechnology Journal (Wiley, CC BY 4.0), Li et al. established a large-scale O-GlcNAc profiling methodology in rice using metabolic glycan labeling with GalNAz combined with click chemistry enrichment, demonstrating the power of bioorthogonal labeling for PTM discovery in non-mammalian systems where antibody-based enrichment reagents are essentially unavailable.
Background: O-GlcNAcylation is a nutrient-sensitive, dynamic modification that regulates transcription, translation, hormone signaling, and stress responses in plants. Despite its recognized importance in plant biology, the O-GlcNAc proteome of rice — the world's most important food crop — remained largely uncharacterized. Conventional O-GlcNAc enrichment using commercial antibodies (e.g., RL2, CTD110.6) suffers from limited specificity, epitope masking, and poor cross-reactivity with plant O-GlcNAc-modified proteins. The authors adopted a bioorthogonal labeling strategy to overcome these limitations.
Approach: Rice seedlings were metabolically labeled with N-azidoacetylgalactosamine (GalNAz), which enters the UDP-GlcNAc salvage pathway and is incorporated by O-GlcNAc transferases (OGTs) into O-GlcNAc-modified Ser/Thr residues. After protein extraction, azide-labeled O-GlcNAcylated proteins were conjugated via CuAAC click chemistry to an alkyne-functionalized biotin tag containing a cleavable linker. Biotinylated proteins were captured on streptavidin agarose, and labeled peptides were released by cleavable linker cleavage. The enriched O-GlcNAc peptides were analyzed by LC-MS/MS with HCD-pd-EThcD fragmentation for unambiguous site localization, as standard HCD alone causes O-GlcNAc neutral loss (HexNAc, +203.08 Da) that precludes site assignment.
Key Findings:
- Scale of O-GlcNAc identification: A total of 1,591 unambiguous O-GlcNAcylation sites were identified on 709 O-GlcNAcylated proteins — representing the largest O-GlcNAc dataset in a plant species at the time of publication, enabled by the bioorthogonal labeling approach that bypassed the coverage limitations of antibody-based enrichment
- Biological scope: Identified O-GlcNAc-modified proteins encompassed transcription factors, RNA-binding proteins, translation machinery components, hormone signaling proteins, and stress-responsive proteins — demonstrating the broad regulatory reach of O-GlcNAcylation across diverse plant cellular processes
- Enzyme discovery: Two functional O-GlcNAc transferases (OsOGT1 and OsOGT2) were identified from the enriched O-GlcNAc proteome and their enzymatic activity was confirmed in vitro, providing the first biochemical characterization of OGT enzymes in rice
- Methodological advantages: The GalNAz metabolic labeling approach identified substantially more O-GlcNAc sites than previous antibody-based studies in plants, and the cleavable linker strategy eliminated streptavidin peptide contamination that commonly compromises LC-MS analysis in workflows using on-bead digestion
- Validation data: Enriched O-GlcNAc peptides showed characteristic HCD-pd-EThcD fragmentation spectra with diagnostic HexNAc oxonium ions (m/z 204.09, 186.08, 168.07, 138.06) and charge-reduced precursor ions, providing high-confidence site localization for >90% of identified O-GlcNAc sites
Significance: This study established metabolic glycan labeling with click chemistry enrichment as a transformative approach for O-GlcNAc proteomics in non-mammalian systems, demonstrating that bioorthogonal labeling can identify modification sites at a scale and confidence level unattainable by antibody-based methods. The workflow — GalNAz metabolic incorporation, CuAAC click chemistry to cleavable biotin, streptavidin enrichment, and HCD-pd-EThcD LC-MS/MS with diagnostic oxonium ion monitoring — is directly transferable to mammalian cells, patient-derived models, and other biological systems where comprehensive O-GlcNAc mapping is required. The dual-enzyme discovery (OsOGT1 and OsOGT2) further demonstrates that bioorthogonal enrichment can identify not only modified proteins but also the modifying enzymes themselves, through their association with labeled substrates during enrichment.
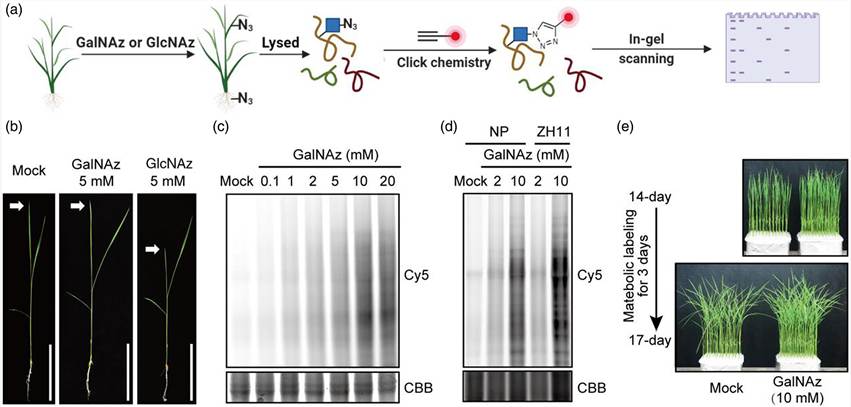
Adapted from Li et al. (2023). Metabolic glycan labeling and click chemistry enrichment for O-GlcNAc proteome profiling in rice. (CC BY 4.0)
Related Services
Our bioorthogonal PTM labeling platform is part of a comprehensive PTM analysis service portfolio spanning chemical labeling, antibody-based enrichment, MS-based discovery, and bioinformatics integration for multi-PTM research programs.
- O-GlcNAc Profiling Services — Dedicated O-GlcNAc analysis using lectin affinity, enzymatic labeling, and MS-based identification for comprehensive O-GlcNAc site mapping in cell and tissue samples
- Protein Lipidation Analysis — Comprehensive MS-based characterization of palmitoylated, myristoylated, and prenylated proteins using chemical proteomics and enrichment strategies
- Glycoproteomics Analysis Services — Integrated glycoproteomics platform combining lectin enrichment, HILIC, and enzymatic deglycosylation for N-glycopeptide and O-glycopeptide identification
- Modified Peptide Enrichment Services — Enrichment solutions including IMAC, TiO₂, HILIC, and antibody-based immunoaffinity approaches for modified peptide isolation prior to MS analysis
- Ultra-Sensitive Modified Protein Detection — Targeted MRM/PRM-based verification of low-abundance modified proteins identified through discovery-phase bioorthogonal labeling experiments
- PTM Bioinformatics Analysis — Advanced bioinformatics analysis of PTM datasets including modification crosstalk, pathway enrichment, motif analysis, and multi-PTM data integration
FAQs
What is the difference between CuAAC and SPAAC click chemistry for bioorthogonal labeling?
CuAAC (copper-catalyzed azide-alkyne cycloaddition) uses a copper(I) catalyst and is faster, more economical, and suitable for lysate-based labeling after cell harvesting. SPAAC (strain-promoted azide-alkyne cycloaddition) uses strained alkynes (DBCO, BCN) and proceeds without copper catalyst, making it suitable for live-cell and in vivo labeling where copper toxicity is a concern. We recommend CuAAC for most cell lysate applications where maximal yield is desired, and SPAAC for live-cell imaging, in vivo labeling in animal models, or applications where copper-induced oxidative stress would confound biological results.
Which bioorthogonal probe should I choose for my PTM of interest?
Probe selection is determined by the target PTM class: GalNAz for O-GlcNAc and O-glycan labeling; ManNAz for sialic acid and sialoglycoproteomics; GlcNAz for N-glycan labeling; alk-C16 (16-alkynyl hexadecanoic acid) for S-palmitoylation; alk-C14 (14-alkynyl tetradecanoic acid) for N-myristoylation; and AHA (azidohomoalanine) for nascent protein and modification turnover labeling. For pilot experiments, we recommend starting with the best-characterized probe for your target modification class and confirming labeling efficiency by in-gel fluorescence before scaling to LC-MS analysis.
How much starting material is required for a bioorthogonal labeling and enrichment experiment?
For cell culture experiments, we recommend starting with 1–5 × 10⁷ cells per condition (approximately 1–5 mg total protein after lysis). For tissue samples, 20–100 mg wet weight is typically sufficient. Probe concentration and labeling time must be optimized for each cell type: typical GalNAz labeling uses 50–200 µM for 24–72 hours, alkynyl fatty acids use 10–100 µM for 4–16 hours, and AHA uses 1–4 mM in methionine-free media for 15–120 minutes. We perform labeling efficiency checks by in-gel fluorescence before committing samples to the full enrichment and LC-MS workflow.
How does cleavable linker technology improve identification compared to on-bead digestion?
On-bead digestion — where trypsin digests proteins while they remain bound to streptavidin beads — releases both labeled target peptides and large amounts of streptavidin, trypsin autolysis products, and bead-derived contaminants that suppress MS signal and consume LC-MS duty cycle. Cleavable linkers (acid-labile, photocleavable, or hydroxylamine-cleavable) enable specific release of labeled peptides under mild conditions, leaving streptavidin and non-labeled contaminants on the beads. This typically increases the number of identified modification sites by 2–5-fold compared to equivalent on-bead digestion workflows.
Can bioorthogonal labeling be performed in vivo in animal models?
Yes. Bioorthogonal probes can be administered to live animals through various routes — intraperitoneal injection, drinking water supplementation, or targeted delivery. GalNAz has been successfully used for in vivo O-GlcNAc labeling in zebrafish and mice, and AHA labeling has been applied for nascent proteome profiling in mice via IP injection. For in vivo applications, SPAAC click chemistry using DBCO-functionalized probes is typically preferred over CuAAC to avoid copper toxicity. Sample collection, tissue homogenization, and click chemistry enrichment are performed after animal sacrifice.
What are the limitations of bioorthogonal labeling that I should consider?
Several limitations should be considered: (1) Metabolic labeling requires live, metabolically active cells — it cannot be applied to archival or fixed tissue samples. (2) Probe incorporation efficiency varies between cell types and may be affected by serum concentration, metabolic flux, and probe toxicity. (3) CuAAC click chemistry requires copper, which can cause protein oxidation and modification artifacts if not properly controlled. (4) The bioorthogonal handle (azide/alkyne) slightly modifies the native substrate structure and may affect incorporation efficiency or biological behavior. (5) Site localization for labile modifications (O-GlcNAc, S-palmitoylation) requires specialized fragmentation methods (EThcD) that may not be available on all MS platforms. We address each of these limitations through optimized protocols, appropriate controls (including no-probe and no-click controls), and access to advanced fragmentation capabilities.
How are bioorthogonal PTM labeling data analyzed differently from conventional PTM proteomics data?
Bioorthogonal labeling data require modification-specific analysis parameters in the database search: O-GlcNAc analysis uses HexNAc as a variable modification (+203.08 Da on Ser/Thr) with diagnostic oxonium ions (m/z 204.09, 186.08) for spectral validation; palmitoylation analysis uses palmitoyl as a variable modification (+238.23 Da on Cys) with hydroxylamine-lability confirmation; myristoylation analysis uses myristoyl as a fixed N-terminal modification (+210.20 Da on N-terminal Gly); and glycoproteomics data require dedicated glycopeptide search engines (Byonic, pGlyco, O-Search) that can match both glycan composition and peptide sequence simultaneously. All identified modification sites are validated by manual spectral inspection with site localization scoring.
References
- Li X, Lei C, Song Q, Bai L, Cheng B, Qin K, Li X, Ma B, Wang B, Zhou W, Chen X, Li J. Chemoproteomic profiling of O-GlcNAcylated proteins and identification of O-GlcNAc transferases in rice. Plant Biotechnology Journal. 2023;21(4):742-753.
- Zaia J. The 2022 Nobel Prize in Chemistry for the development of click chemistry and bioorthogonal chemistry. Analytical and Bioanalytical Chemistry. 2023;415(4):527-532.
- Wu R, Ji G, Chen W, Zhang L, Fang C, Lu H. Simultaneous and site-specific profiling of heterogeneity and turnover in protein S-acylation by intact S-acylated peptide analysis with a cleavable bioorthogonal tag. Analyst. 2024;149(4):1111-1120.
For research use only. Not for use in diagnostic procedures.