
Protein glycosylation is a process whereby oligosaccharides covalently bind to specific amino acid residues on proteins in the form of glycosides. Protein glycosylation can be divided into four categories based on the connection method between the amino acid and the sugar: O-linked glycosylation, N-linked glycosylation, C-mannosylation, and glycosylphosphatidylinositol (GPI)-anchoring.
O-glycosylation predominantly occurs on serine or threonine residues proximal to proline, most proteins at the glycosylation site are beta conformers. O-linked oligosaccharides are formed by stepwise addition of monosaccharides to form oligosaccharides. So far, no specific protein sequence has been found as a glycosylation site. O-glycosylation reactions occur at two locations within the cell, one at the Golgi apparatus and the other in the nucleus or cytoplasm.
Glycosylation occurring in the Golgi apparatus initiates by adding N-acetylglucosamine, N-acetylgalactosamine, mannose, fucose, and other reducing ends to the hydroxyl group of serine and threonine. O-glycosylation of secreted proteins and membrane-bound proteins happens after N-glycosylation and protein folding, it occurs on the cis face of Golgi.
Glycosylation taking place in the nucleus and cytoplasm involves adding a single sugar, N-acetylglucosamine, to serine or threonine residues. The most common form of O-glycosylation in mammals is O-GalNAc glycosylation catalyzed by GalNAc transferase, which subsequently attaches Gal, GalNAc, or GlcNAc moieties.
Select Service
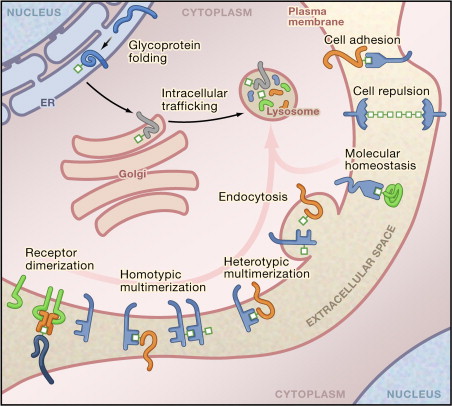 Glycosylation in Cellular Mechanisms of Health and Disease (Kazuaki Ohtsubo et al,. 2006)
Glycosylation in Cellular Mechanisms of Health and Disease (Kazuaki Ohtsubo et al,. 2006)
Regarding the O-GlcNAc glycosylation, it is divided into two conformations: O-α-GlcNAc and O-β-GlcNAc, with the glycosylation process being reversible. The glycosylation is characterized by three main features: the glycosylation site is similar to the protein kinase action site; glycosylation and phosphorylation mutually inhibit each other, with O-GlcNAc glycosylation having a dephosphorylation effect on numerous proteins; and the dynamics of O-GlcNAc glycosylation allow it to respond rapidly to cellular signals, among other functions.
In the process of N-linked glycosylation, a sugar moiety is catalytically attached to the asparagine (Asn) amino acid residue of endocrine proteins and membrane-bound proteins within the endoplasmic reticulum. It is generally accepted that N-linked glycosylation takes place on the sequenced protein Asn-Xaa-Ser/Thr, where Xaa represents all amino acid residues except proline. In rare cases, the Asn-Xaa-Cys sequence also acts as a glycosylation site.
C-mannosylation involves the attachment of an α-mannopyranosyl moiety to the C-2 position of indole ring of tryptophan residue via a C-C bond. This glycosylation event predominantly occurs on the first tryptophan residue of the W-X-X-W, W-X-X-C, or W-X-X-F motif. Glycosylphosphatidylinositol (GPI) anchoring, on the other hand, entails the fusion of phosphatidylinositol and glycan moiety near the C-terminus of a protein, serving as a tether to the cell membrane.
Analysis of glycosylation sites
The determination of glycosylation sites requires the digestion and separation of proteins, followed by detection methods such as mass spectrometry (MS) or tandem mass spectrometry (MS/MS). The combination of liquid chromatography (LC) and MS provides a reliable method for identifying glycosylation sites in hydrolyzed mixtures. Krokhin and others have utilized MALDI technology to identify and determine glycosylation sites, and are embarking on the development of automated identification software for glycopeptide precursor ions.
Synthesis of Glycoproteins
Synthesis of glycoproteins has been a persistent impediment in the study of the impacts of protein glycosylation on protein function and structure, especially when selective modifications on analogs are involved, thus rendering chemical synthesis considerably challenging. In 2004, Zhang et al. reported a co-translational synthesis method for obtaining selectively glycosylated modified proteins. The modified amino acids can be encoded by genes. This method can be widely applied to the synthesis of proteins with other forms of post-modifications. Concurrently, it also provides a potential avenue for the preparation of glycopeptide drugs. Recent research has identified an N-glycosylation pathway in human gastrointestinal pathogen Campylobacter jejuni, as well as an O-glycosylation pathway within C. jejuni and its closely related species Campylobacter coli. These findings present model systems for the study of glycopeptide biosynthesis. The N- and O-glycosylation pathways in the aforementioned bacterial systems are analogous with those in eukaryotes, featuring similar biological functions. Homologs of their genes can also be found in other organisms. Polysaccharide complexes linked to glycopeptides share common biosynthetic precursors.
The Role of Glycosylation in Biological Systems
Protein glycosylation, influencing its function, plays an important function in numerous biological processes such as immune protection, viral replication, cell growth, cell-to-cell adhesion, and the progression of inflammation. Many proteins, such as transcription factors, nucleoporins, heat shock proteins, RNA polymerase II, oncoprotein translational products, and enzymes, have shown post-translational modifications in the form of glycosylation. Glycosylation abnormalities often result in disease onset. For instance, elevated levels of transferrin glycosylation are observed in patients suffering from Parkinson's disease, rheumatoid arthritis, and other diseases associated with free radicals. Transferrin, a glycosylated metal transporter serum protein, provides stability to transferrin glycosylation and indirectly regulates iron balance. Extensive literature has associated glycosylation dysregulation with rapidly developing muscular dystrophies: for example, Muntoni, et al. identified a novel mechanism leading to muscle disorders through the identification of glycosyltransferase activity. In all the cases studied, non-normal α-dystroglycan glycosylation was universally present.
Glycosylation by Li Yanmei et al. investigated the synthesis of functionally modified peptides with O-GlcNAc modification, the impact of O-GlcNAc modification on peptide conformation and biological function, and the relationship and impact of O-GlcNAc and O-phosphorylation modification regulation. Given the significant role of glycosylation in organisms, drugs that target glycosylation sites have emerged in succession with therapeutic mechanisms involving the regulation of the corresponding enzymes.
References
- Blom N, Sicheritz-Pontén T, Gupta R, et al. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics, 2004, (4): 1633~1649
- Hart G W. Dynamic O-linked glycosylation of nuclear and cytoskeletal proteins. Annu Rev Biochem, 1997, 66 315~335
- Asker N, Baeckstrom D, Axelsson M, et al. The human MUC2 mucin apoprotein appears to dimerize before O-glycosylation and shares epitopes with the 'insolub 33 le' mucin of rat small intestine. 34 ral 35 increases toward the C terminus. Protein 36 eptides. Rapid Commun 37 e Biochem J, 1995, 308: 873~880
- Roth J, Wang Y, Eckhardt A E, et al. Differential expression of cell surface sialoglycoconjugates on wild-type and cultured ehrlich tumor cells as revealed by quantitative lectin-gold ultrastructu cytochemistry. Proc Natl Acad Sci USA, 1994, 91: 8935~8939
- Gavel Y, von Heijne G. Statistical studies of N-glycosylated proteins have indicated that the frequency of nonglycosylated AsnXaa-(Thr/Ser) sequons Eng, 1990, 3: 433~442
- Krokhin O, Ens W, Standing K G, et al. Site-specific N-glycosylation analysis: matrix-assisted laser desorption/ionization quadrupole-quadrupole time-of-flight tandem mass spectral signatures for recognition and identification of glycop Mass Spectrom, 2004, (18): 2020~2030
- Zhang Z W, Gildersleeve J, Yang Y Y, et al. A new strategy for th synthesis of glycoproteins. Science, 2004, 303: 371~373
- Szymanski C M, Logan S M, Linton D, et al. Campylobacter a tale of two protein glyc 38 osylation systems. Trends in Microbiology, 39 tion in cell signaling. Science, 1995, 268: 40 l of Biological Chemistry, 2000, 275(38): 2003, 11(5): 233~238
- Cassey P J. Protein lopida 221~225 Comer F I , Hart G. W. O-Glycosylation of Nuclear and Cytosolic Proteins. The Journa 29179~29182
- van Rensburg S J, Berman P, Potocnik F, et al. 5-and 6-glycosylation of transferrin in patients with Alzheim 41 er's disease. Metabolic Brain Disease, 2004, 19(1-2): 89~96
- Muntoni F, Brockington M, Torelli S, et al. Defective glycosylation in congenital muscular d 42 ystrophies. Current Opinion in Neurology, 2004, 17 (2): 205~209
- Dwek R A, Butters T D, Platt F M, et al. Targeting glycosylation as a therapeutic app 43 roach. Nature Reviews Drug Discovery, 2002, 1(1): 65~75
.jpg)
Download


Our products and services are for research use only.
